FÍSICA BÁSICA DE LOS POTENCIALES DE MEMBRANA
Potenciales de membrana provocados por difusión
Potencial de Nernst: Es el nivel del potencial de difusión a través de una membrana que se opone exactamente a la difusión neta de un ion particular a través de la membrana.
Cuando una membrana es permeable a varios iones diferentes, el potencial de difusión que se genera depende de tres factores:
1) la polaridad de la carga eléctrica de cada uno de los iones.
2) la permeabilidad de la membrana a cada uno de los iones.
3) las concentraciones de los respectivos iones en el interior y en el exterior de la membrana. Para ello se usa una fórmula denominada ecuación de Goldman, que proporciona el potencial de membrana calculado en el interior de la membrana cuando participan dos iones positivos univalentes.
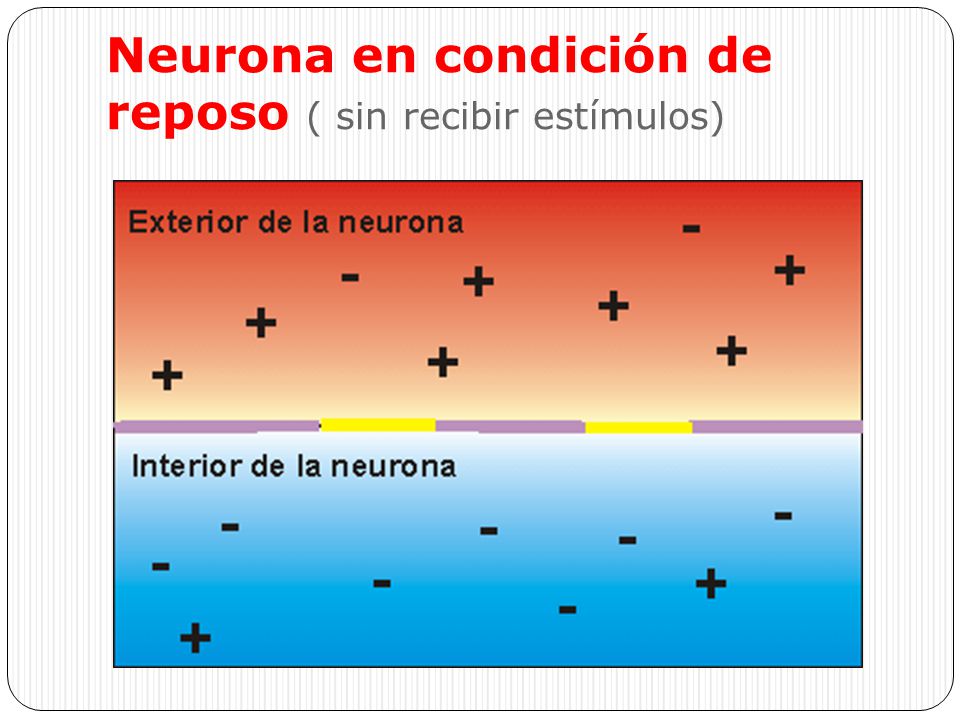
Potencial de membrana en reposo de los nervios
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es alrededor de –90 mV .
Bomba Na-K
Todas las membranas celulares del cuerpo tienen una potente bomba Na+K+ que transporta continuamente iones sodio hacia el exterior de la célula e iones potasio hacia el interior; ésta bomba también es conocida como bomba electrógena porque se bombean más cargas positivas hacia afuera de la célula que hacia el adentro (3 Na hacia afuera por cada 2 K hacia adentro), dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular
Fuga de potasio y de sodio a través de la membrana nerviosa.
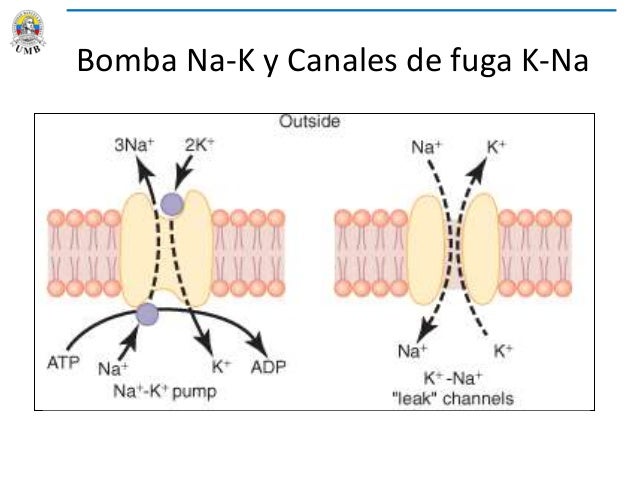 Una proteína del canal, a veces denominada dominio de poros en tándem, canal de potasio o canal de fuga de potasio también puede dejar que se pierdan algunos iones de sodio, sin embargo, siempre van a ser más permeables al K; ésta diferencia de permeabilidad es un factor importante para determinar el nivel del potencial de membrana en reposo normal.
Una proteína del canal, a veces denominada dominio de poros en tándem, canal de potasio o canal de fuga de potasio también puede dejar que se pierdan algunos iones de sodio, sin embargo, siempre van a ser más permeables al K; ésta diferencia de permeabilidad es un factor importante para determinar el nivel del potencial de membrana en reposo normal.
Potencial de membrana en reposo normal
el potencial de membrana en reposo normal de –90 mV, y los factores que lo determinan son:
♦ potencial de difusión de potasio.
♦ potencial de difusión de sodio.
Éstos dos factores se hacen posible gracias a los canales de fuga del K, y como se mencionó anteriormente, éstos canales permiten una mínima difusión de Na, por lo que es lógico pensar que la difusión del potasio contribuye mucho más al potencial de membrana que la difusión del sodio.
♦ Contribución de la bomba Na+K+.
Por lo tanto, los potenciales de difusión aislados que produce la difusión del sodio y del potasio dan un potencial de membrana de aproximadamente –86 mV, casi todo determinado por la difusión de potasio. Además, se generan –4 mV adicionales al potencial de membrana por la acción continua de la bomba de Na+K+ electrógena, generándose un potencial neto de membrana de –90 mV.
Potencial de acción nervioso
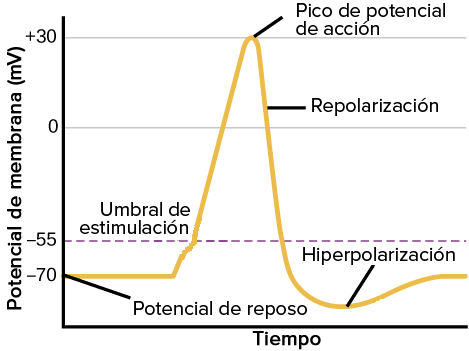
 Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito o inesperado desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y después termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para poder conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega al extremo de la misma.
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito o inesperado desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y después termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para poder conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega al extremo de la misma.
Las fases del potencial de acción:
 Fase de reposo: Es el potencial de membrana en reposo antes del comienzo del potencial de acción, la membrana está polarizada durante esta fase debido al potencial de membrana negativo de –90 mV que posee.
Fase de reposo: Es el potencial de membrana en reposo antes del comienzo del potencial de acción, la membrana está polarizada durante esta fase debido al potencial de membrana negativo de –90 mV que posee.
Fase de despolarización: En este momento la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado polarizado normal de –90 mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva. Esto se denomina despolarización. 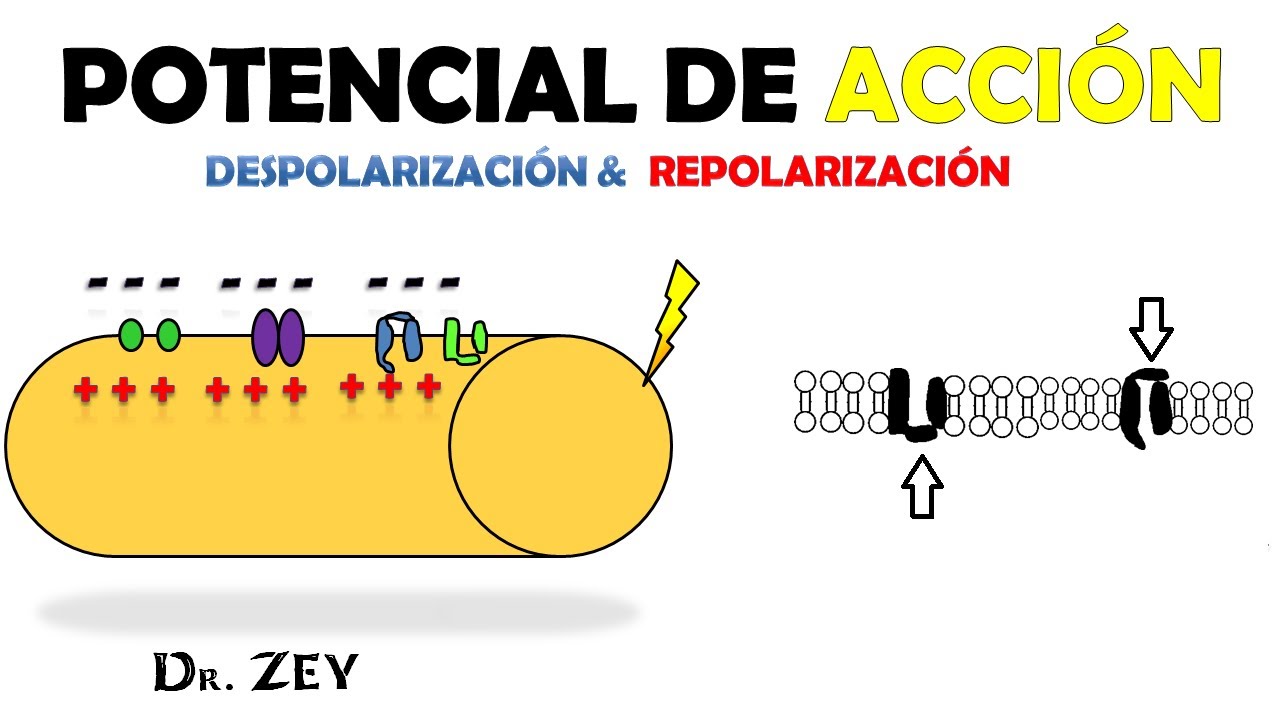
Fase de repolarización: después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal.
Los canales de sodio y potasio activados por el voltaje tienen una función adicional a la
de la bomba Na+K+ y de los canales de fuga K+.
Canal de sodio: tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación.
 Activación: Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje ( entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta pudiendo así pasar los iones sodio.
Activación: Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje ( entre –70 y –50 mV) que produce un cambio conformacional súbito en la activación de la compuerta pudiendo así pasar los iones sodio.
Desactivación: El mismo aumento de voltaje que abre la compuerta de activación también cierra la compuerta de inactivación, después de que el canal de sodio haya permanecido abierto se cierra la compuerta de inactivación y los iones sodio ya no pueden pasar hacia el interior de la membrana.
Un dato importante es que el canal de sodio no se puede abrir de nuevo sin que antes se repolarice la fibra nerviosa. Los canales de sodio se pueden bloquear por una toxina denominada tetrodotoxina aplicándola al exterior de la membrana celular en la que están localizadas las compuertas de activación del sodio.
Canal de potasio:
 Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90 mV hacia cero, este voltaje produce que la compuerta se abra y permite el aumento de la difusión de potasio hacia fuera a través del canal, pero debido a que dilata un poco la apertura de los canales de potasio, en su mayor
Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90 mV hacia cero, este voltaje produce que la compuerta se abra y permite el aumento de la difusión de potasio hacia fuera a través del canal, pero debido a que dilata un poco la apertura de los canales de potasio, en su mayor
parte, se abren al mismo tiempo que se están cerrando los canales de sodio debido a su inactivación.
El ion tetraetilamonio bloquea los canales de potasio cuando se aplica al interior de la fibra nerviosa
La disminución de la entrada de sodio hacia la célula y el aumento simultáneo de la salida de potasio de la célula se combinan para acelerar el proceso de repolarización, lo que da lugar a la recuperación completa del potencial de membrana en reposo en diezmilésimas de segundo.
Pinza de voltaje
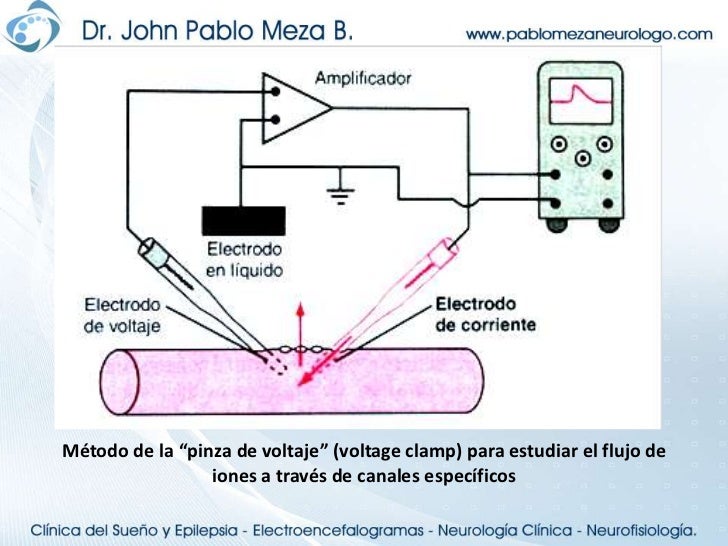 Es un método experimental que se utiliza para medir el flujo de iones a través de los diferentes canales. Cuando se utiliza este aparato se insertan dos electrodos en la fibra nerviosa. Uno de ellos sirve para medir el voltaje del potencial de membrana y el otro para conducir corriente eléctrica hacia el interior o el exterior de la fibra nerviosa.
Es un método experimental que se utiliza para medir el flujo de iones a través de los diferentes canales. Cuando se utiliza este aparato se insertan dos electrodos en la fibra nerviosa. Uno de ellos sirve para medir el voltaje del potencial de membrana y el otro para conducir corriente eléctrica hacia el interior o el exterior de la fibra nerviosa.
Funciones de otros iones durante el potencial de acción
Aniones no difusibles en el interior del axón nervioso: Estos iones no pueden salir del interior del axón, por lo que cualquier deficiencia de iones positivos en el interior de la membrana deja un exceso de estos aniones negativos no difusibles. Por tanto, éstos son responsables de la carga negativa en el interior de la fibra cuando hay un déficit neto de iones potasio de carga positiva y de otros iones positivos.
Iones calcio: Las membranas de casi todas las células del cuerpo tienen una bomba de calcio similar a la bomba de sodio, y el calcio coopera con el sodio (o actúa en su lugar) en algunas células para producir la mayor parte del potencial de acción. Al igual que la bomba de sodio, la bomba de potasio transporta iones calcio desde el interior hacia el exterior de la membrana celular.  Una función importante de los canales de iones calcio activados por voltaje consiste en su contribución a la fase de despolarización en el potencial de acción en algunas células. Se denominan canales lentos, en contraposición a los canales de sodio, que se denominan canales rápidos. Por tanto, la apertura de los canales de calcio proporciona una despolarización más sostenida, mientras que los canales de sodio desempeñan un papel clave en la iniciación de los potenciales de acción. Hay abundantes canales de calcio tanto en el músculo cardíaco como el músculo liso.
Una función importante de los canales de iones calcio activados por voltaje consiste en su contribución a la fase de despolarización en el potencial de acción en algunas células. Se denominan canales lentos, en contraposición a los canales de sodio, que se denominan canales rápidos. Por tanto, la apertura de los canales de calcio proporciona una despolarización más sostenida, mientras que los canales de sodio desempeñan un papel clave en la iniciación de los potenciales de acción. Hay abundantes canales de calcio tanto en el músculo cardíaco como el músculo liso.
Inicio del potencial de acción
Si un estímulo produce una elevación suficiente del potencial de membrana desde –90 mV hacia el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos canales de sodio activados por el voltaje, lo que permite que iones sodio se adentren en la célula rapidamente elevando el potencial de membrana. Posteriormente, en un plazo diminuto, el aumento del potencial de membrana produce cierre de los canales de sodio y apertura de los canales de potasio, y pronto finaliza el potencial de acción. 
Umbral
 No se puede producir un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen. Esto se produce cuando el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Se dice que el nivel de –65 mV es el umbral para la estimulación.
No se puede producir un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen. Esto se produce cuando el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Se dice que el nivel de –65 mV es el umbral para la estimulación.
Propagación del potencial de acción

Un potencial de acción que se desencadena en cualquier punto de una membrana excitable habitualmente excita porciones adyacentes de la membrana, dando lugar a la propagación del potencial de acción a lo largo de la membrana. Esta transmisión del proceso de despolarización a lo largo de una fibra nerviosa muscular se denomina impulso nervioso o muscular. 
Dirección de la propagación.
Una membrana excitable no tiene una dirección de propagación única, sino que el potencial de acción viaja en todas las direcciones alejándose del estímulo hasta que se ha despolarizado toda la membrana.
Principio del todo o nada.
Una vez que se ha originado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización viaja por toda la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no lo son. Esto se denomina principio del todo o nada.
Metabolismo de la energía
Con el tiempo se hace necesario restablecer las diferencias de las concentraciones de membrana de sodio y de potasio. Esto se consigue por la acción de la bomba Na+K+. Los iones sodio que han difundido hacia el interior de la célula durante los potenciales de acción y los iones potasio que han difundido hacia el exterior deben volver a su estado original por la bomba Na+K+. Como esta bomba precisa energía para esta operación, esta recarga de la fibra nerviosa es un proceso metabólico activo que utiliza la energía que procede del sistema energético del trifosfato de adenosina (ATP) de la célula. Una característica especial de esta bomba es que su grado de actividad se estimula mucho cuando se acumula un exceso de iones sodio en el interior de la membrana celular.
Meseta en algunos potenciales de acción
 En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y sólo después comienza la repolarización. Este tipo de potencial de acción se produce en las fibras musculares cardíacas.
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y sólo después comienza la repolarización. Este tipo de potencial de acción se produce en las fibras musculares cardíacas.
La causa de la meseta es una combinación de varios factores. En primer lugar, en el proceso de despolarización del músculo cardíaco participan dos tipos de canales:
1) los canales de sodio (canales rápidos).
2) los canales de calcio-sodio (canales lentos).
La apertura de los canales rápidos origina la porción en espiga del potencial de acción, mientras que la apertura prolongada de los canales lentos de calcio-sodio principalmente permite la entrada de iones calcio en la fibra, lo que es responsable en buena medida también de la porción de meseta del potencial de acción.
Un segundo factor es que los canales de potasio activados por el voltaje tienen una apertura más lenta de lo habitual.
Descarga repetitiva
Las descargas repetitivas autoinducidas aparecen normalmente en el corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso central. Estas descargas rítmicas producen:
1) el latido rítmico del corazón.
2) el peristaltismo rítmico de los intestinos.
3) fenómenos neuronales, como el control rítmico de la respiración.



Para que se produzca ritmicidad espontánea la membrana, incluso en su estado natural, debe ser lo suficientemente permeable a los iones sodio (o a los iones calcio y sodio a través de los canales lentos de calcio-sodio) como para permitir la despolarización automática de la membrana.
Características especiales de la transmisión de señales en los troncos nerviosos
Fibras nerviosas mielinizadas y no mielinizadas.
 El núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de mielina, que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier.
El núcleo central de la fibra es el axón, y la membrana del axón es la membrana que realmente conduce el potencial de acción. El axón contiene en su centro el axoplasma, que es un líquido intracelular viscoso. Alrededor del axón hay una vaina de mielina, que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier.
Las células de Schwan le propporcionan mielina al axón, En la unión entre dos células de Schwann sucesivas a lo largo del axón permanece una pequeña zona no aislada de sólo 2 a 3 mm de longitud en la que los iones pueden seguir fluyendo con facilidad a través de la membrana del axón entre el líquido extracelular y el líquido intracelular del interior del axón. Esta zona se denomina nódulo de Ranvier.
Conducción saltatoria en las fibras mielinizadas de un nódulo a otro.
 Los iones no pueden fluir a través de las gruesas vainas de mielina de los nervios mielinizados, pueden fluir fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen sólo en los nódulos. Esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro.
Los iones no pueden fluir a través de las gruesas vainas de mielina de los nervios mielinizados, pueden fluir fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen sólo en los nódulos. Esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro.
La conducción saltatoria es útil por dos motivos:
1.- Al hacer que el proceso de despolarización salte intervalos largos a lo largo del eje de la fibra nerviosa, este mecanismo aumenta la velocidad de la transmisión nerviosa en las fibras mielinizadas.
2.- La conducción saltatoria conserva la energía para el axón porque sólo se despolarizan los nódulos.
Otra característica adicional de la conducción saltatoria en las fibras mielinizadas gruesas es el excelente aislamiento que ofrece la membrana de mielina y la disminución de 50 veces de la capacitancia de la membrana permiten que se produzca la repolarización con poca transferencia de iones
Velocidad de conducción en las fibras nerviosas.
 La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan sólo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan sólo 0,25 m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
Excitación: el proceso de generación del potencial de acción
Un potencial de acción se puede deber a un trastorno mecánico de la membrana, a los efectos químicos o al paso de electricidad a través de ella. Todos ellos se utilizan en diferentes puntos del cuerpo para generar potenciales de acción nerviosos o musculares.
Excitación de una fibra nerviosa por un electrodo metálico cargado negativamente.
El método habitual para excitar un nervio o un músculo en el laboratorio experimental es aplicar electricidad a la superficie del nervio del músculo mediante dos electrodos pequeños, uno de los cuales tiene carga negativa y el otro positiva.
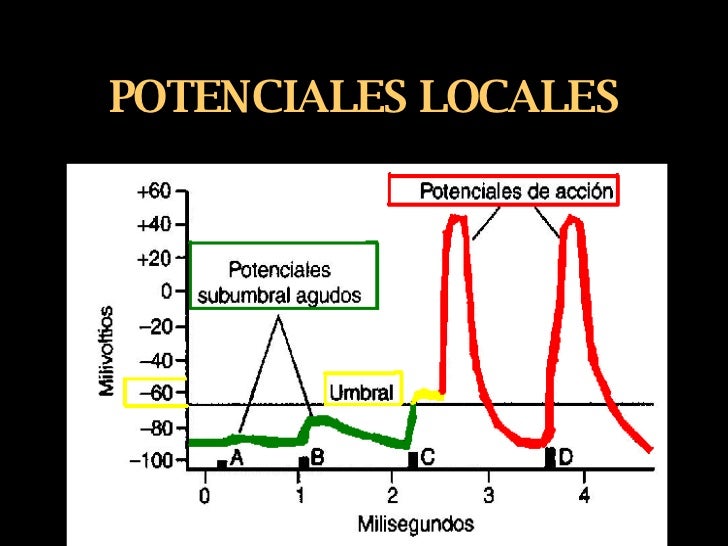 Los cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generarun potencial de acción, se denominan potenciales subliminales agudos. Cuando el potencial local apenas alcanza el nivel necesario para generar un potencial de acción, se denomina nivel liminar (umbral).
Los cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generarun potencial de acción, se denominan potenciales subliminales agudos. Cuando el potencial local apenas alcanza el nivel necesario para generar un potencial de acción, se denomina nivel liminar (umbral).
Estabilizadores y anestésicos locales
 Los factores estabilizadores de la membrana son aquellos que pueden reducir la excitabilidad, por ejemplo, el ion calcio es un estabilizador.
Los factores estabilizadores de la membrana son aquellos que pueden reducir la excitabilidad, por ejemplo, el ion calcio es un estabilizador.
Anestésicos locales: procaína y tetracaína
Registro de potenciales de membrana y potenciales de acción
Osciloscopio de rayos catódicos.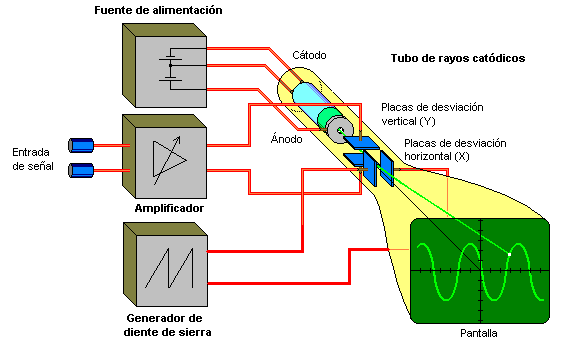
 La membrana celular está formada en su mayoría por una bicapa lipídica, aunque también contiene varias moléculas proteicas insertadas en los lípidos, muchas de ellas penetran en todo el grosor de la membrana.
La membrana celular está formada en su mayoría por una bicapa lipídica, aunque también contiene varias moléculas proteicas insertadas en los lípidos, muchas de ellas penetran en todo el grosor de la membrana.  Otras, denominadas
Otras, denominadas 
 Entre las sustancias más importantes que atraviesan las membranas celulares mediante difusión facilitada están la
Entre las sustancias más importantes que atraviesan las membranas celulares mediante difusión facilitada están la  Por ejemplo, los poros proteicos denomi-nados
Por ejemplo, los poros proteicos denomi-nados Muchos de los canales proteicos son muy selectivos para el transporte de uno o más iones o moléculas específicos. Esto se debe a las características del propio canal, como su diámetro, su forma y la naturaleza de las cargas eléctricas y enlaces químicos que están situados a lo largo de sus superficies internas.
Muchos de los canales proteicos son muy selectivos para el transporte de uno o más iones o moléculas específicos. Esto se debe a las características del propio canal, como su diámetro, su forma y la naturaleza de las cargas eléctricas y enlaces químicos que están situados a lo largo de sus superficies internas. 

























 El aparato locomotor contribuye a la homeostasis cuando permite la movilidad del individuo cuando éste se desplaza en busca de sus alimentos.
El aparato locomotor contribuye a la homeostasis cuando permite la movilidad del individuo cuando éste se desplaza en busca de sus alimentos.
 El sistema nervioso central está formado por el cerebro y la médula espinal. El cerebro almacena información, genera los pensamientos, y determina las reacciones que debe manifestar el cuerpo en respuesta a las sensaciones para, a continuación, transmitir las señales apropiadas a través de la porción motora eferente del sistema nervioso para llevar a cabo los deseos del sujeto.
El sistema nervioso central está formado por el cerebro y la médula espinal. El cerebro almacena información, genera los pensamientos, y determina las reacciones que debe manifestar el cuerpo en respuesta a las sensaciones para, a continuación, transmitir las señales apropiadas a través de la porción motora eferente del sistema nervioso para llevar a cabo los deseos del sujeto. 










 Este aparato es prominente en las células secretoras, dondemse localiza en el lado de la célula a partir del cual se extruirán las sustancias secretoras.
Este aparato es prominente en las células secretoras, dondemse localiza en el lado de la célula a partir del cual se extruirán las sustancias secretoras.